国立研究開発法人 産業技術総合研究所【理事長 中鉢 良治】(以下「産総研」という)生物プロセス研究部門【研究部門長 田村 具博】 深津 武馬 首席研究員(兼)生物共生進化機構研究グループ 研究グループ長、細川 貴弘 元産総研特別研究員(現 九州大学 助教)らは、放送大学、琉球大学、沖縄科学技術大学院大学と協力して、腸内共生細菌が生存に必須な農業害虫のチャバネアオカメムシにおいて、北海道、本州、四国、九州等(以下「日本本土」という)の集団では共生細菌は単一種であるが、南西諸島では同じ集団中に複数種の共生細菌が共存することを発見した。
南西諸島のカメムシ集団を詳細に解析したところ、(1)カメムシ体外では培養できず、垂直感染だけで次世代に伝達される共生細菌、(2) カメムシ体外で培養可能で環境中でも生存でき、垂直感染も環境獲得もおこり、他種カメムシにも感染可能な共生細菌、(3) 通常はカメムシに共生しないが、実験的に感染させると潜在的な共生能力を示す環境細菌、という進化段階の異なるさまざまな共生細菌が共存していた。すなわち南西諸島においては、チャバネアオカメムシの腸内共生関係は日本本土のように確立しておらず、今もダイナミックな進化過程にあることがわかった。
従来、新しい高度な共生関係が成立する際の具体的な過程や機構はよくわかっていなかったが、現在進行中の共生進化の過程を自然界でとらえたことで、共生の起源と進化の理解が大きく進展するとともに、害虫の環境適応機構の理解とその制御への展開も期待される。
この成果は2016年1月11日23時(日本時間)に国際科学雑誌Nature Microbiology(ネイチャー・マイクロバイオロジー)にオンライン掲載される。
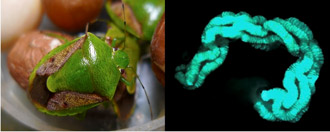 |
チャバネアオカメムシ(左)の中腸共生器官(右)
摘出した消化管内の青緑色は局在する腸内共生細菌を示す |
ヒトを含む多種多様な動物の消化管内には、多様な細菌、真菌、原生生物などから構成される腸内微生物叢が存在し、さまざまな生物機能を担っている。近年、ヒトでも健康、病気、肥満などに腸内細菌叢が関与するという報告が相次ぎ、腸内微生物叢の多様性、機能、起源に関する研究は大きな展開をみせている。
カメムシ類は世界で4万種以上、日本だけでも1,500種余が知られ、農作物の害虫であることも多い。このため、農業や経済の観点から、その生物機能の詳細な解明が待たれる重要な昆虫群であるが、消化管の後端部が共生器官として特殊化し、成長や繁殖に必須な特定の(しばしば単一種の)細菌を保有する。これらの細菌はビタミンや必須アミノ酸などの栄養素を供給しており、多くの場合カメムシの体外では増殖できない。
このように高度な生物機能を担う特殊化した共生細菌も、もともとは環境中に存在する普通の細菌であったと考えられるが、どのような過程と仕組みにより、環境細菌から高度な共生細菌が生じてきたのかについては、よくわかっていなかった。
産総研では、これまでに昆虫の体内に共生する細菌が有する重要な生物機能の解明や、昆虫と共生細菌の間の高度な生物間相互作用の理解などに取り組んできた。特に農業害虫として重要なカメムシ類の腸内共生細菌については、「共生細菌による昆虫の害虫化の発見」(2007年6月13日 産総研プレス発表)、「害虫に殺虫剤抵抗性を持たせる共生細菌を発見」(2012年4月24日 産総研プレス発表)、「昆虫と細菌との共生におけるポリエステルの新たな機能」(2013年6月11日 産総研プレス発表)、「害虫カメムシが共生細菌を体内に取り込む特異な仕組みを解明」(2015年9月1日 産総研プレス発表)などの成果をあげてきた。今回、農業害虫であるチャバネアオカメムシについて、日本列島における腸内共生細菌の多様性を発見し、その形成過程と機構の解明に取り組んだ。
なお本研究は、独立行政法人 日本学術振興会の科学研究費助成事業・基盤研究(S)の支援を受けておこなった。
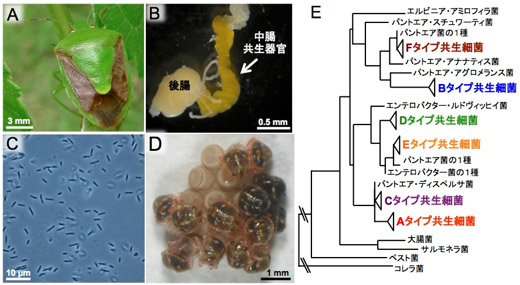
チャバネアオカメムシ(図1A)の消化管の後部には特殊化した共生器官があり(図1B)、その内部に大腸菌と近縁の共生細菌(ここでは共生細菌Aとよぶ)を保有している(図1C)。この共生細菌は幼虫の成長に不可欠で、母虫は産卵の際に共生細菌入りの分泌物を卵の表面に塗布し、孵化した幼虫がそれを摂取することで共生細菌が垂直感染する(図1D)ことがわかっていた。
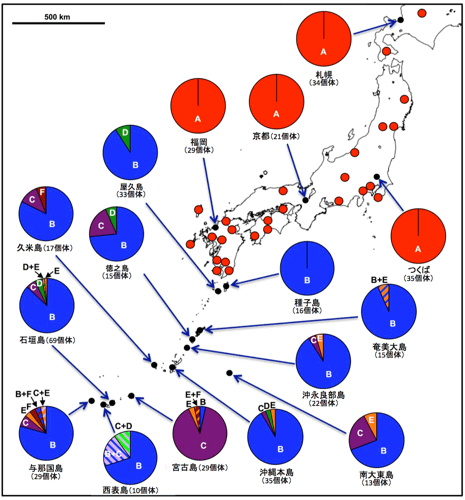
従来は本州のチャバネアオカメムシが研究の対象であったが、今回、日本列島全域から収集した42集団448個体について腸内共生細菌を調べたところ、6種類の異なる腸内共生細菌(ここでは共生細菌A、B、C、D、E、Fとよぶ)が同定された(図1E)。これらのうち、日本本土の集団では共生細菌Aだけが検出されたが、南西諸島の集団では共生細菌Bが高頻度で検出される一方で、共生細菌C、D、E、Fが低頻度で共存していた(図2)。なお、南西諸島の中でも宮古島では、例外的に共生細菌Cが高頻度で検出された。
 |
|
図1 チャバネアオカメムシと腸内共生細菌 |
|
(A) 成虫、(B) 摘出した消化管後部(共生器官を矢印で示す)、(C) 共生細菌の光学顕微鏡像、(D) 卵殻表面から共生細菌を獲得中の孵化幼虫、(E) 日本の集団が保有する共生細菌6種の系統関係(リボソームRNA遺伝子の塩基配列に基づく) |
 |
|
図2 日本のチャバネアオカメムシ集団の腸内共生細菌の感染分布 |
卵表面を滅菌して孵化幼虫への共生細菌の垂直感染を妨げて除去したところ、共生細菌を除去した虫は、ほとんどが幼虫期に死亡し、かろうじて成虫になった個体は矮小で異常な体色を呈していた(図3)。同様の結果が共生細菌A、B、C、D、E、Fに共通して得られ、6種の共生細菌がいずれも幼虫の正常な成長に必須であることがわかった。
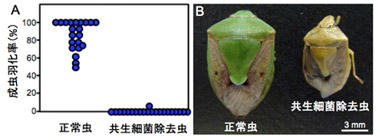 |
図3 腸内共生細菌除去の宿主カメムシへの影響
(A) 成虫羽化率、(B) 羽化してきた成虫 |
また、共生器官をすりつぶして培養を試みたところ、共生細菌A、Bはまったく増殖しなかったが、共生細菌C、D、E、Fは容易に培養できた。概要ゲノム配列を決定したところ、共生細菌A、Bのゲノムは共生細菌C、D、E、Fのゲノムに比べて小さく、おそらく共生が進化する過程でゲノム縮小が進んだため宿主体外での増殖が困難になったと考えられる。
さらに、もともと共生細菌Bに感染していたチャバネアオカメムシの卵表面を滅菌して、孵化した幼虫に培養した共生細菌C、D、E、Fを与えて共生細菌を置き換えたところ(図4A)、もとの共生細菌に感染したものと同等の生育を示した(図4B)。もともと共生細菌Aに感染していたチャバネアオカメムシを用いても同様の結果が得られ、これら6種の共生細菌は宿主の成長に必須な同様の生理機能を担っていると考えられる。
 |
|
図4 腸内共生細菌置換の宿主カメムシへの影響 |
|
(A) 培養共生細菌の実験感染システム、(B) もとの共生細菌Bに感染したカメムシ(左端)と同様に、共生細菌C、D、E、Fに実験的に置換したカメムシ(右側)も正常に成長する |
共生細菌C、D、E、Fが培養可能であるということは、自然環境中で生存している可能性がある。そこで表面を滅菌した卵から生まれたチャバネアオカメムシの孵化幼虫を、石垣島の生息地から採取した土壌に6日間曝露(図5A)してから、清浄な餌と容器で飼育したところ、計1,005匹の孵化幼虫の大部分は死亡したが、85匹が成虫まで到達した。うち14匹は矮小で異常な体色を呈し、共生細菌を保有しないと考えられたが(図5C)、残りの71匹は正常な成虫であった(図5B、D-L)。うち過半数は共生細菌C、D、Eのいずれかに感染しており、残りは新規な細菌(仮に環境細菌X1-X6とよぶ)に感染していた(図5B-L)。これらの結果から、環境中にはチャバネアオカメムシの培養可能な共生細菌C、D、Eが存在すること、また多様な環境細菌の中には潜在的にチャバネアオカメムシへの共生能力をもつものが含まれることが明らかになった。
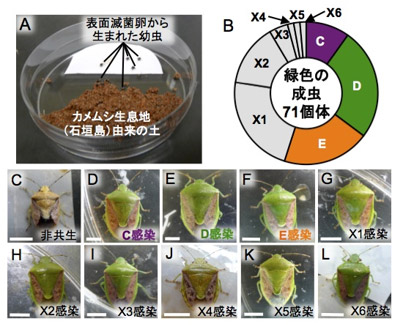 |
図5 土壌環境からの共生細菌獲得の実験的証明
(A) 土壌細菌の実験感染システム、(B) 緑色の羽化成虫に感染していた細菌の内訳、(C-L) 羽化成虫の外観と感染していた細菌 |
南西諸島に分布する多種多様なカメムシ類について調べたところ、チャバネアオカメムシの共生細菌C、D、Eと遺伝子配列レベルで区別がつかない細菌を腸内に保有する種が少なからずいた(図6A-C)。そこで、もともと共生細菌Bに感染していたチャバネアオカメムシの卵表面を滅菌して、孵化幼虫にこれら他種カメムシの共生器官から分離培養した共生細菌C、D、Eを与えて、共生細菌を置き換えたところ、もとの共生細菌Bに感染したものと同等の生育を示した(図6D)。すなわち、環境中に存在する共生細菌C、D、Eはおそらく、チャバネアオカメムシに限らずさまざまなカメムシ類とも共生関係にあり、母虫から幼虫へ垂直感染するだけではなく、時に環境中から獲得されたり、あるいは他種カメムシから水平感染するなど、カメムシ群集と環境細菌群集の間にダイナミックな関係が成立している可能性が考えられる。
 |
|
図6 他種カメムシとの間の腸内共生細菌置換の影響 |
|
(A-C) チャバネアオカメムシ腸内共生細菌C、D、Eと遺伝的に区別できない共生細菌を保有する他種カメムシ、(D) もとの共生細菌Bに感染したカメムシ(左端)と同様に、他種由来の共生細菌C、D、Eに置換したカメムシも正常に成長する。 |
このように、日本列島ではチャバネアオカメムシの腸内共生細菌叢がダイナミックな進化過程にある。日本本土では、宿主体外では生存できない共生細菌Aがカメムシ集団中に完全に固定しているのに対し、南西諸島ではやはり宿主体外では生存できない共生細菌Bがカメムシ集団中に完全に固定するには至らず、宿主体外でも生存でき、環境細菌のような性質をもち、他種カメムシにも共生できる共生細菌C、D、E、Fも共存している。さらに、カメムシへの共生能力を潜在的にもつさまざまな環境細菌が土壌環境中に存在することもわかった。これらの研究により、現在進行中の共生進化の過程を自然界でとらえることに成功し、共生関係の起源および進化過程の理解が大きく進展した。南西諸島は日本列島でも特筆すべき豊かな生物多様性を擁するが、本研究でその生物多様性のこれまで知られていなかった新しい側面を解明した。
今後は、南西諸島における共生細菌の多様性を野外のチャバネアオカメムシ集団で経時的にモニターするとともに、実験室においてさまざまな環境条件で継続飼育した場合の共生関係への影響を調べる実験進化的な研究も推進する。特に、日本本土では1種の共生細菌に固定する一方で、なぜ南西諸島では多様な共生細菌が共存しているのかを解明する。
さらに、共生細菌間の競争や共存などの相互作用について解析を進め、共生機構の共通性と多様性について理解を深めるとともに、農業害虫であるカメムシ類の新規防除標的として、共生細菌を利用する可能性を検討する。特に温帯気候の日本本土で感染が固定した共生細菌Aと、亜熱帯気候の南西諸島で感染が優占する共生細菌Bは、異なる温度条件や寄主植物への適応に関わっている可能性があり、そのような観点からの研究も展開していく予定である。