独立行政法人 産業技術総合研究所【理事長 野間口 有】(以下「産総研」という)バイオメディカル研究部門【研究部門長 織田 雅直】RNAプロセシング研究グループ 富田 耕造 研究グループ長らは、鋳型非依存性RNA合成酵素のうち、mRNAの末端へポリA配列を合成付加する真正細菌由来のポリA付加酵素の構造を決定し、基質特異性を規定する分子機構を明らかにした。
細胞内のmRNAは通常、DNAから転写された後、その末端に、DNA上の配列とは無関係に数十から数百のアデニンヌクレオチド(ポリA配列)がポリA付加酵素と呼ばれる鋳型非依存性RNA合成酵素によって付加される。mRNAのポリA配列は、RNAの安定性や、翻訳などに関わっており、遺伝子発現に重要な役割を果たしている。なお、ポリA付加酵素を含む多種多様な鋳型非依存性RNA合成酵素群が、欠陥RNAを積極的に分解して除去するRNA品質管理や多岐にわたる遺伝子発現に関与するノンコーディングRNAの発現制御などに関わっていることが知られている。しかしながら、ポリA付加酵素の基質に対する特異性の分子レベルでのメカニズムは不明であった。本研究により、RNA合成酵素のデザイン開発が加速され、鋳型非依存性RNA合成酵素の反応制御を解明することで新たな医薬開発への応用が期待される。本成果は、米国Cell出版社のStructure誌に掲載された。
 |
|
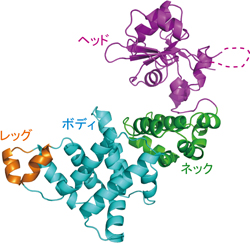
ポリA付加酵素の全体構造:ヘッド、ネック、ボディ、レッグからなるラッコ型構造をとる
|
細胞内における遺伝情報はDNA→mRNA→タンパク質という順に伝わっている。つまり、DNA鎖を鋳型としてmRNAへ遺伝情報が転写される。その後、mRNAは多種多様な加工を受け、最終的に機能しうるmRNAへと成熟化される。mRNAへの転写後の加工過程のひとつとして、その末端に鋳型に依存しないRNA合成酵素のひとつであるポリA付加酵素によって数十から数百のアデニンヌクレオチド(ポリA配列)が付加されることが知られている。
mRNAの末端に合成付加されたポリA配列は、そのRNAの安定性や翻訳の促進などに関わっており、遺伝子発現に重要な役割を担っている。また、最近では、生体内における鋳型非依存性RNA合成酵素によるRNA末端へのRNA合成付加が機能欠陥RNAの分解に積極的に関与していることも報告されてきている。すなわち、鋳型非依存性RNA合成酵素は新たなRNAの品質を管理する役割をも有しているといえる。また、この鋳型非依存性RNA合成酵素が、遺伝子制御に深く関わっているタンパク質のアミノ酸配列情報をもっていない低分子ノンコーディングRNAなどの発現制御に関与していることも報告されてきている。したがって鋳型非依存性RNA合成酵素群の詳細な反応のメカニズム、そしてその酵素活性が生体内のどこで、どのタイミングで現れるかを解明することは、新たなRNA合成酵素のデザインのみならず、新たな遺伝子発現の制御解明においても重要な役割を担っている。
産総研では、RNAを合成する酵素群の機能と構造に関する研究を行っている。これまで、鋳型を用いないでRNAを合成する酵素、すなわち鋳型非依存性RNA合成酵素群やウイルス由来の鋳型を用いてRNAを合成する酵素の研究を行ってきている(プレス発表など:2006年10月16日、2008年7月8日、2009年10月5日、2010年8月24日)。本研究は鋳型を用いないRNAの合成酵素のうち、これまで産総研で解析を行ってきたCCA付加酵素とアミノ酸配列がよく似ているが異なった活性を有するポリA付加酵素を対象として、X線結晶構造解析および機能解析を行った。CCA付加酵素、ポリA付加酵素を含む、鋳型非依存性RNA合成酵素群は、多種多様な遺伝子発現に関与している。したがって、これらの鋳型非依存性RNA合成酵素の基質認識の分子機構や反応機構の解明、さらにはその反応制御機構の解明をすることによって、それらの酵素の反応を生体内のどこでどのタイミングで発現させるかを制御することが可能となり新たな医薬品の開発の基盤を提供しうると考えられる。
本研究は、独立行政法人 科学技術振興機構(JST)戦略的創造研究推進事業個人型研究(さきがけ)「RNAと生体機能」【研究総括:野本 明男 微生物化学研究所長】の一環として行われた。また、独立行政法人 日本学術振興会(JSPS)基盤研究Aおよび、財団法人東レ科学振興財団からの研究補助金によって行われた。
細胞内のmRNAはDNAから転写された後、その末端に、DNA上の配列とは無関係にポリA配列がポリA付加酵素と呼ばれる鋳型非依存性RNA合成酵素によって合成付加される。mRNAのポリA配列は、RNAの安定性や、そのmRNAの翻訳促進などに関わっており、遺伝子発現に重要な役割を果たしている。また、最近ではこれらの鋳型非依存性RNA合成酵素が、多岐にわたる遺伝子発現に関与していることが知られてきている。
興味深いことに、真正細菌のポリA付加酵素はtRNAの末端に普遍的に存在するCCA配列を鋳型非依存的に合成するCCA付加酵素とアミノ酸配列がよく似ており、アミノ酸配列のみからではそれらの活性を予測することが難しい。本研究では真正細菌のポリA付加酵素の特異性をもたらす分子レベルでの仕組みや分子レベルでのCCA付加酵素の特異性との違いを明らかにすることを目的として、X線結晶構造解析および生化学的解析を行った。
 |
|
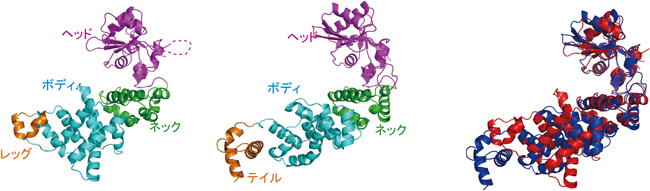
図1 真正細菌ポリA付加酵素の構造(左;ラッコ型)とCCA付加酵素(中央;タツノオトシゴ型)の構造、ポリA付加酵素(赤)とCCA付加酵素(青)の重ね合わせ。ヘッド、ネックドメインの構造はよく似ている(右)。
|
今回のX線結晶構造解析および生化学的解析から、以下のことが明らかになった。
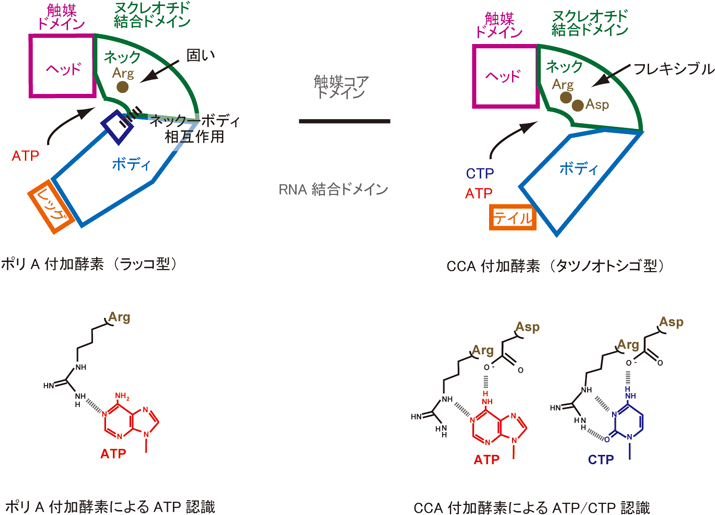
1)ポリA付加酵素の全体構造は、ヘッド、ネック、ボディ、レッグの4つのドメインからなるラッコのような構造をとっており、タツノオトシゴのようなCCA付加酵素の構造とは異なっていた(図1左、中央)。
2)しかしながら、ポリA付加酵素の活性触媒部位やヌクレオチド結合部位を含むヘッド、ネックドメイン構造は、CCA付加酵素のそれらの構造と非常によく似ていた(図1右)。
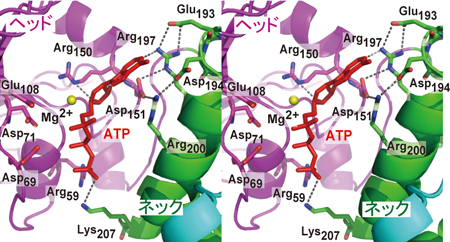
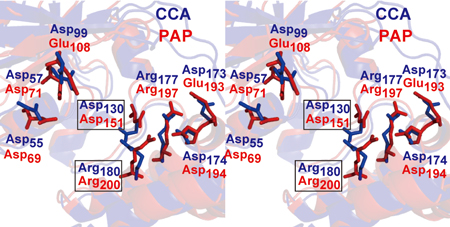
3)ポリA付加酵素のネックドメインに存在するヌクレオチド結合ポケット内の塩基認識に関わると考えられるアミノ酸は分子内水素結合によって固い構造をとっていた(図2)。
4)ポリA付加酵素はネックドメインに存在する固いポケット内で、ひとつの水素結合によってATPを認識しており、そのポケットの形と大きさはATPのみに適したものであった。
5)一方、CCA付加酵素ではヌクレオチドの結合ポケットの塩基認識に関わるアミノ酸は、ポリA付加酵素とは異なり、分子内水素結合を形成しておらず、フレキシブルな構造をとっており、それによってCTPもATPも同時に認識できる(図3)。
6)ポリA付加酵素のヌクレオチドとの結合ポケットの固さは、ネックドメインと、RNA認識に関わるボディドメインとの相互作用に起因することが示唆された(図1、図4)。
7)また、ポリA付加酵素のレッグドメインはポリAが付加されたRNAが酵素を移動するために必要な領域であることが明らかになった。
 |
|
図2 ポリA付加酵素によるATP(赤)の認識(ステレオ図):ATP結合ポケット内のアミノ酸同士の水素結合(灰色破線)によって固い構造をとっている。
|
 |
|
図3 ポリA付加酵素(赤:PAP)、CCA付加酵素(青:CCA)のヌクレオチド結合ポケットの重ね合わせ(ステレオ図)。四角で囲ったアミノ酸の立体構造が二つの酵素では異なる。
|
 |
|
図4 ポリA付加酵素(左)とCCA付加酵素(右)のヌクレオチド認識の違い
|
これらの解析から、永年不明であった、分子レベルでのポリA付加酵素とCCA付加酵素との特異性の差異について明らかになった。
近年、生体内には多種多様な鋳型非依存性RNA合成酵素が存在し、生体内でのRNAの代謝に深く関わっていることが報告されてきている。今後、多種多様な遺伝子発現に関与する他の種類の鋳型非依存性RNA合成酵素についても注目し、それらの酵素の詳細な分子レベルでの反応のメカニズムや遺伝子発現制御における役割の解明を目指したい。
独立行政法人 産業技術総合研究所
バイオメディカル研究部門 RNAプロセシング研究グループ
研究グループ長 富田 耕造 E-mail: