独立行政法人 産業技術総合研究所【理事長 野間口 有】(以下「産総研」という)生物機能工学研究部門【研究部門長 織田 雅直】機能性核酸研究グループ 富田 耕造 研究グループ長、董 雪松 産総研特別研究員、竹下 大二郎 専門技術者(財団法人 日本産業技術振興協会)沼田 倫征 研究員らは、真正細菌由来の鋳型非依存性RNA合成酵素である「CCA付加酵素」が正しい長さのRNAを合成する分子機構を提唱した。
細胞内でタンパク質が合成される際、必要なアミノ酸を運んでくるtRNAという70-90ヌクレオチドの1本鎖RNAがある。このtRNAはDNAを鋳型として合成されるが、tRNAの末端にあるCCAという配列だけはCCA付加酵素という特別な酵素によって、DNAの鋳型を用いずに合成される。これまで産総研は古細菌のCCA付加酵素の反応について分子機構の解明を行ってきた。
今回、真正細菌のCCA付加酵素の解析により、古細菌のCCA付加酵素の場合と同様、酵素タンパク質とRNAが共同で機能を発揮しているが、古細菌の場合とは異なった分子機構で反応が進行することを見いだした。
本研究により、RNA合成酵素のデザイン開発が加速され、バイオテクノロジーへの応用が期待される。本成果は、2009年9月10日(英国時間)に、EMBO Journalにオンライン掲載された。
 |
|
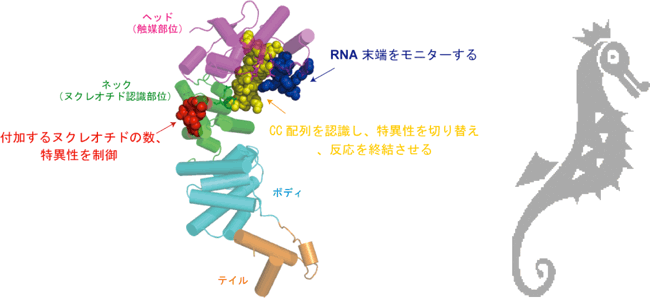
タツノオトシゴ形状のCCA付加酵素によるRNA合成伸長、終結を規定する分子機構
(棒と糸は酵素タンパク質を、赤青黄の球は合成を制御する酵素内の領域を表す。)
|
細胞内における遺伝情報はDNA→mRNA→タンパク質という順に伝わっている。mRNAの配列はDNA配列のコピーであり、その鋳型となるDNAをもとに鋳型依存的なRNA合成酵素によって合成される。一方、生体内には鋳型を用いることなく、定まった配列を合成する鋳型非依存的RNA合成酵素が存在する。鋳型非依存的RNA合成酵素は遺伝子発現に重要な役割を果たしていることが知られているが、詳細な反応の分子機構にはいまだに明らかにされていない点が多い。
近年、生体内における鋳型非依存的RNA合成酵素による鋳型非依存的なRNA付加が、不要になったRNAの分解に積極的に関与していることが報告され、RNAの品質を管理する役割をもっていると考えられるようになってきた。そのため、鋳型非依存的RNA合成酵素の詳細な反応機構の研究は、新たな遺伝子発現制御の解明において重要な役割を担っている。
私たちはこの数年、鋳型非依存的なRNA合成酵素であるCCA付加酵素について一連のX線結晶構造解析、生化学的解析による機構解明を行ってきた。古細菌のCCA付加酵素については、30年間謎であった反応の分子機構、正しい配列を合成する忠実性維持機構を解明してきた(2006年10月16日、2008年7月8日 プレス発表)。真正細菌、真核生物由来のCCA付加酵素は、アミノ酸配列が古細菌由来の物と大きく異なり、その反応分子機構はいまだに解明されていない。
今回、真正細菌由来のCCA付加酵素の反応分子機構解明を目指し、X線結晶構造解析、構造を基にした遺伝学的、生化学的解析を行った。
本研究は、独立行政法人 科学技術振興機構 戦略的創造研究推進事業個人型研究(さきがけ)「RNAと生体機能」【研究総括 野本 明男】の課題として、また財団法人 東レ科学振興財団からの研究補助金によって行われた。
次のような一連の試料についてX線結晶構造解析、生化学的解析、遺伝学的解析を行った。
|
1) |
真正細菌由来のCCA付加酵素のみ |
|
2) |
真正細菌由来のCCA付加酵素とCTP(反応原料のC塩基)複合体 |
|
3) |
真正細菌由来のCCA付加酵素とATP(反応原料のA塩基)複合体 |
|
4) |
真正細菌にはCCA付加酵素の他にA塩基のみを付加するA付加酵素が存在するが、この2つの酵素はよく似た形状で、ヘッド(頭)、ネック(首)、ボディー(体)、テイル(尾っぽ)の4つの部分から構成されている。遺伝子操作によりA付加酵素のヘッド(頭)やネック(首)部分をCCA付加酵素のそれらの部分と置き換えた酵素(キメラ酵素という)を作製し、比較検討した。 |
その結果、以下のことが明らかになった。
|
1) |
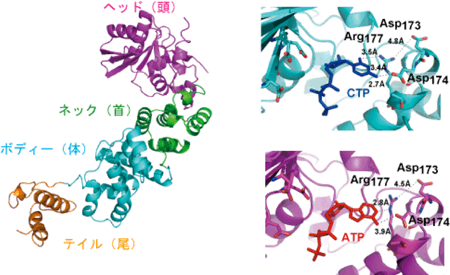
真正細菌のCCA付加酵素はタツノオトシゴに似た形をとり、ヘッド(頭)、ネック(首)、ボディー(体)、テイル(尾っぽ)の4つの部分からなる。ヘッド部分は活性触媒部位を含み、ネック部分はヌクレオチドの認識にかかわっている。 |
|
2) |
ATPとCTPはヘッドとネックの間のくぼみに入っており、ネック部分の同じアミノ酸によって認識されている。
 |
|
図2 真正細菌CCA付加酵素の全体構造。タツノオトシゴに似た形(左)をとり、CTPとATPは同じポケット(くぼみ)によって認識されている(右)。(右上図はCTPがポケットに入った様子、右下図はATPがポケットに入った様子)
|
|
|
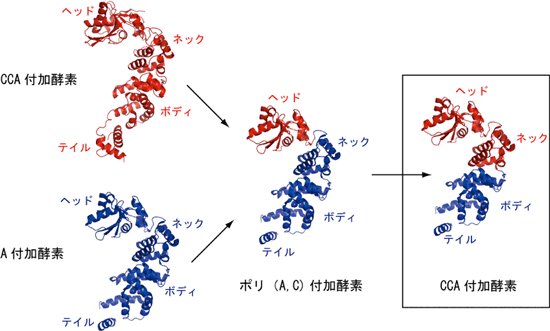
3) |
さまざまなキメラ酵素の解析から、A付加酵素のヘッド部分をCCA付加酵素のヘッド部に置き換えただけではCCA付加酵素にならないが、ヘッドとネック部分両方を置き換えるとCCA付加酵素になることがわかった。ヘッド部分だけを交換したときにはCとAがランダムに付加されてしまい、また、合成されるRNA鎖の長さもいろいろになり、反応の終結を制御できなかった。
 |
|
図3 CCA付加酵素(赤)とA付加酵素(青)のキメラ酵素の解析。ヘッドとネックの両方の交換によってA付加酵素がCCA付加酵素へと変換された。
|
|
|
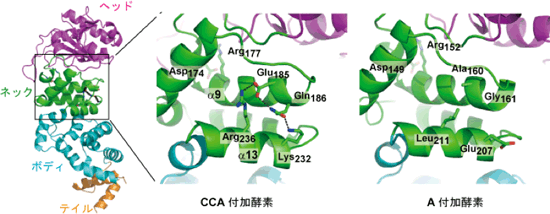
4) |
ネック部分内の3本のヘリックス間の水素結合が、ヌクレオチドの特異性および付加されるヌクレオチドの数を規定するばね状のちょうつがいの役割を果たし、ランダムなCとAの付加反応を抑制している。
 |
|
図4 ネック内のヘリックス間の水素結合がばね状のちょうつがいの役割をはたし、付加されるヌクレオチドの数、特異性を規定している。
|
|
|
5) |
ヘッド部分内のループ領域が最後のA付加反応に関与していることが判明した。タンパク質のループ領域が、伸長してきたRNA末端のCC配列を認識すると、ヘッドとネック間のポケット(くぼみ)の構造が変化してATPのみが入れるようになり、最後のAを付加して合成は終結する。
 |
|
図5 ヘッド内のループ(黄色)によるCC配列認識によるポケット(くぼみ)の構造と最後のATPの認識モデル。右図は左図の表面モデル。
|
|
以上のような解析により私たちは真正細菌のCCA付加酵素が、正しいRNA鎖を合成する次のような分子機構を提唱した。この酵素によるCCA合成反応は、ヘッド部分とネック部分の動的かつ共同的な作用で進行する。この共同的な作用において、ネック部分がばね状のちょうつがいの役割を果たし、付加されるヌクレオチドの数、特異性を規定していることが示唆された。また、RNAのCC配列が合成されると、その配列をヘッド部分のループが認識して、ポケットの構造を変化させてAのみを特異的に取り込むようになり、A付加反応が進行し反応は終結する。
この研究で明らかになった真正細菌CCA付加酵素によるRNA合成伸長、終結分子機構はこれまで解析が行われた古細菌のそれとは異なった分子機構である。しかしながら、RNAとタンパク質が共同で機能を発現してという点は共通している。
現在、真正細菌由来のCCA付加酵素がCCA配列を合成する過程のすべての複合体の構造解析や、他の鋳型非依存的RNA合成酵素の解析をも進めている。鋳型非依存的なRNA合成酵素の詳細な分子機構の解明、分子進化、他の種類の鋳型非依存性RNA合成酵素についても解析をすすめ、RNAとタンパク質が共同で機能を発揮する詳細な分子機構解明、さらに新機能RNA合成酵素の創生を目指す。