国立研究開発法人 産業技術総合研究所【理事長 石村 和彦】(以下「産総研」という)生物プロセス研究部門【研究部門長 鈴木 馨】 微生物生態工学研究グループ 菊池 義智 研究グループ付 (兼)国立大学法人 北海道大学 大学院農学院 客員准教授、伊藤 英臣 主任研究員らは、国立大学法人 北海道大学【総長 寳金 清博】(以下「北大」という) 大学院農学院 博士課程学生 Jang Seonghan、石神 広太と共同で、国立研究開発法人 農業・食品産業技術総合研究機構【理事長 久間 和生】、大学共同利用機関法人 自然科学研究機構 基礎生物学研究所【所長 阿形 清和】、フランス国立科学研究センター(CNRS)と協力し、昆虫の呼吸器官である「気管」の形成時に、活性酸素種が気管を構成するタンパク質の架橋に関与し、その硬化と形態維持に重要な役割を果たすことを明らかにした。
昆虫は肺を持たず、身体中に張り巡らされた気管によって呼吸をする。気管は、節足動物が陸上に進出するにあたり獲得した新たな呼吸器官であり、その形成メカニズムの解明は昆虫の進化を紐解く上で学術的に重要であるばかりでなく、新たな発想の害虫防除技術の開発にも繋がりうる。今回、気管の硬化と形態維持に活性酸素種が重要な役割を果たすことを明らかにした。また、昆虫の腸内に生息する好気性細菌が、酸素を消費することで気管の形成を促進することも明らかにした。これら気管形成に関わるメカニズムの発見は、気管の形成を阻害するような新しい発想の防除技術の開発につながる可能性がある。
なお、この研究の詳細は、米国の学術誌「Proceedings of the National Academy of Sciences USA」(米国科学アカデミー紀要)に近日中にオンライン掲載される。
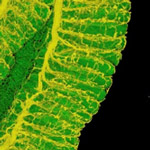
ホソヘリカメムシの消化管(緑)を取り巻くように複雑に発達する気管(黄色)
昆虫は肺を持たず、「気管」で酸素と二酸化炭素を直接交換して呼吸する。気管はクチクラで裏打ちされたチューブ状の構造で、昆虫の体全体に張り巡らされ、体表面の開口部(気門)に通じている。気管は、それまで水中でエラ呼吸を行っていた昆虫の祖先が、陸上に進出し空気呼吸を効率的に行うために進化させた器官であり、これによって陸上でも効率的に酸素を利用でき、飛翔のような俊敏でダイナミックな動きができるようになったと考えられている。気管は昆虫の陸上適応に重要な役割を果たしてきたと考えられるが、その形成メカニズム、特にその頑健性をもたらす硬化のメカニズムについてはほとんど解明されていなかった。昆虫に特有の生理生態、さらにそれをもたらす遺伝的基盤や生化学的メカニズムの解明は、学術的に重要である。
産総研は、害虫の新たな生理生態、多くの害虫が持つ共生微生物の機能の解明を目指して研究を行ってきた。特にこれまで、ダイズを食害する農業害虫のホソヘリカメムシをモデル系として研究を進め、その生理生態や腸内共生細菌の機能についての研究に取り組んできた (2015年9月1日、2018年1月18日産総研プレス発表)。この過程で、昆虫腸内細菌の多様性や機能を明らかにし、遺伝子組換え技術や昆虫の遺伝子抑制技術(RNAi)を確立し、共生現象を多面的に解析する技術を高度化してきた。一方、北大は産総研との連携大学院を設けており、環境中の微生物に関する研究を進めてきた。
今回、両者はホソヘリカメムシが腸内に共生する好気性細菌をどのように維持しているのか、そのメカニズムを研究する過程で、気管形成に関わるメカニズムを発見した。
なお、本研究は、日本学術振興会科学研究費助成事業(15H05638,19K15724)、特別研究員奨励費(201911493)による支援を受けて行った。
Dual oxidase (Duox)は活性酸素種を産生する酵素として知られている。いくつかの昆虫では、消化管の上皮細胞で発現するDuoxによって産生された活性酸素種が消化管内腔に放出され、腸内細菌の抑制に重要な役割を果たしていることが報告されているが、腸内共生細菌の維持にDuoxが果たす役割については不明な点が多い。ホソヘリカメムシの腸内共生細菌は消化管に発達した袋状組織(盲嚢)に共生している。今回、RNAiによってホソヘリカメムシのDuoxの発現を抑制したところ、共生細菌の量が著しく減少していた(図1)。
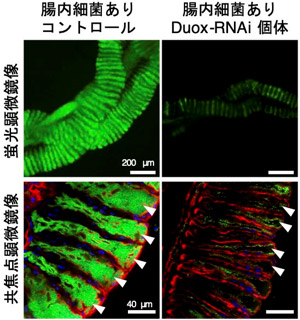
図1 Duoxを発現抑制すると腸内共生細菌が減少
緑は腸内共生細菌、共焦点顕微鏡像の青はホソヘリカメムシの核、赤は細胞骨格を示す。
ホソヘリカメムシ消化管のDuox発現部位を調べたところ、Duoxは消化管の細胞で発現しているのではなく、消化管を取り巻く気管で強く発現していた(図2)。これまでに、Duoxによって生成される活性酸素種がタンパク質中のチロシンを架橋し、ジチロシン結合(DTN)を形成することで組織の形態維持や硬化にも重要な役割を果たすことが知られていた。そこで、ホソヘリカメムシ消化管をジチロシン抗体で免疫染色し観察したところ、ジチロシンが気管で特異的にみられた。また、DuoxをRNAiにより阻害したり、抗酸化物質であるN-アセチルシステインをホソヘリカメムシに与えたりして、活性酸素種の生成を抑制するとジチロシン形成、つまり気管形成が著しく阻害されることが分かった(図2)。
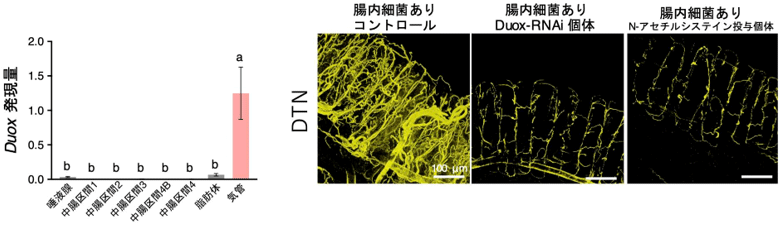
図2 気管におけるDuoxの発現とその機能
左は各組織におけるDuoxの発現量、右は腸管のジチロシン結合(DTN)の検出。
左の棒グラフで、Duox発現量に統計的有意差がみられた組織を異なるアルファベットで示す。
共生細菌は好気性なので、気管の形成不全により消化管内腔への酸素供給が滞って共生細菌の増殖が阻害され減少した可能性が考えられる。実際に、Duoxの発現を抑制した個体の共生細菌は酸素欠乏時に特徴的な遺伝子発現パターンを示していた。また、Duoxの発現抑制を行い、気管形成を阻害した個体を酸素濃度40%のチャンバー内で飼育すると、気管の形成が阻害されているにもかかわらず共生細菌の量は大きく増加した(図3)。これらの結果から、Duoxは気管の形成に重要な役割を果たし、これによって腸内の酸素濃度が増加し、共生細菌が安定的に維持されるものと考えられる。
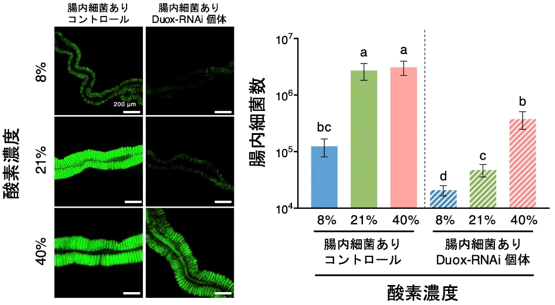
図3 酸素濃度が腸内細菌量に及ぼす影響
左は腸内の共生細菌を緑色蛍光で観察した像、右は定量PCRによる腸内細菌量の測定値。
統計的有意差がみられた実験区を異なるアルファベットで示す。
また、共生細菌に感染していない個体では気管の発達が著しく低いことが分かった(図4)。これは、腸内に定着した共生細菌が気管の形成を誘導していることを示している。おそらく腸内の共生細菌に酸素を効率よく供給するために、腸の気管形成が活性化されると考えられる(図5)。また、RNAiによって気管の形成誘導に関わる転写因子(Bnl, Trh, Sima)の発現を抑制すると、Duoxの発現抑制時と同様に、気管の形成が著しく阻害され(図4)、同時に共生細菌量も大きく減少した。
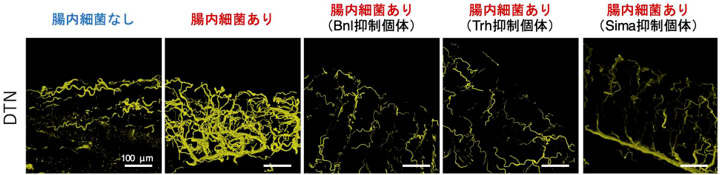
図4 ホソヘリカメムシの気管形成状態
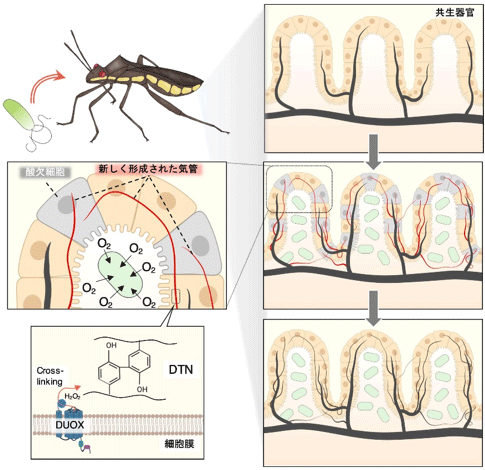
図5 腸内細菌による気管形成の誘導とDuoxによる気管硬化メカニズムの概略
この現象がホソヘリカメムシに特異的な現象であるどうかかを確かめるために、モデル昆虫であるコオロギ、カイコ、コクヌストモドキについてDuox発現量を計測してみたところ、中腸組織よりも気管での発現が非常に高かった。さらに、ショウジョウバエの遺伝子発現データベースを調査したところ、ショウジョウバエでもDuoxの気管での発現量が高かった(図6)。これらは、Duoxによる気管の硬化・形態形成は昆虫に広く見られる一般的な現象であることを強く示唆している。
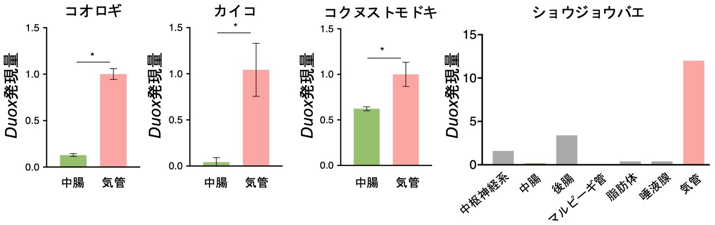
図6 各種昆虫のDuoxの発現量
*は統計的有意差があることを示す。
今後は、昆虫以外にも気管を発達させているクモ類のような陸生節足動物の活性酸素種産生酵素(Duox)の機能について解析を進め、節足動物が陸上環境に進出する際に、どのような進化が起きたのかを調べる。また、今回の発見は害虫の気管形成を阻害する新たな防除技術の開発につながるかもしれない可能性があることから、Duoxの機能を阻害する化合物や抗酸化物質の研究開発も展開していく。