国立研究開発法人 産業技術総合研究所【理事長 中鉢 良治】(以下「産総研」という)生物プロセス研究部門【研究部門長 田村 具博】 植物機能制御研究グループ 坂本 真吾 研究員、光田 展隆 研究グループ長らは、メルボルン大学、ブリティッシュコロンビア大学などと共同して、新しく同定した一次細胞壁形成を制御する遺伝子を使い、従来の木質(二次細胞壁)のかわりにリグニンがなく極めて酵素糖化性の高い細胞壁を高蓄積させることに成功した。
植物の細胞壁は、どの細胞にも普遍的な一次細胞壁と、強度を必要とする細胞(道管や繊維細胞)に蓄積する二次細胞壁(木質)に大別できる。二次細胞壁に多く含まれるリグニンはバイオマス分解を阻害し、燃料や物質生産の障害となる。今回、リグニンを含まない一次細胞壁の形成を制御する遺伝子としてERF転写因子群を発見した。これを、二次細胞壁を形成しないシロイヌナズナ変異体に導入することで、リグニンがなく酵素糖化性の高い一次細胞壁に似た細胞壁を繊維細胞に高蓄積させることに成功した。今回発見した転写因子を利用した木質改変植物の開発により、木質バイオマスを利用する工程で必要なエネルギーや化学薬品を減らすことができ、二酸化炭素排出削減への貢献が期待される。さらに今回発見した遺伝子の機能解析を進めることで、いまだ多くが解明されていない植物の一次細胞壁の形成機構が明らかになり、植物の「植物らしさ」を解明することにつながると期待される。
なお、この技術開発の詳細は、2018年10月1日(現地時間)にNature Plantsにオンライン掲載される。
 |
|
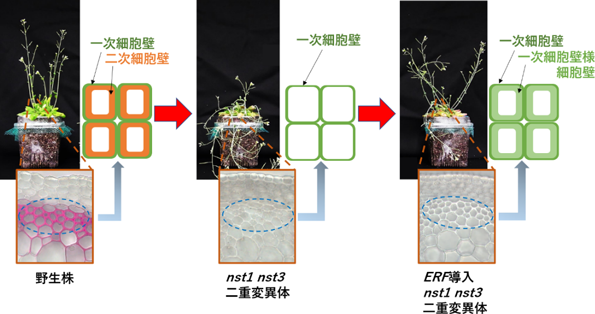
シロイヌナズナと細胞壁の顕微鏡像 |
地球温暖化の主要な原因となる二酸化炭素の放出を抑制するため、植物などの生物資源を材料にした物質生産や燃料生産が注目を集めている。特に食糧生産と競合しない木質バイオマス資源は、次世代バイオ燃料や次世代バイオ素材であるセルロースナノファイバーの供給源として注目されている。しかし、木質バイオマスに大量に含まれるリグニンはバイオマス分解を阻害するため、その先の工程である物質生産や燃料生産の障害となる。リグニンの除去には大きなエネルギーや化学薬品の投入が必要であるため、リグニンの少ない木質バイオマス資源の開発が求められていた。
産総研では、工業原料・医薬品・食料生産などへの応用を目指し、植物転写因子に関する研究開発を進めてきた。特に植物細胞壁の研究では、植物の二次細胞壁(木質)の形成を制御するNST転写因子群の発見、イネNST転写因子を利用した「木質増強ポプラ」の開発など、植物細胞壁の研究で世界をリードする研究成果を挙げてきた。
今回は、産業利用しやすい木質原料を生産する植物の研究開発に取り組んだ。なお、この研究は、国立研究開発法人 科学技術振興機構(JST) 戦略的創造研究推事業 先端的低炭素化技術開発(ALCA)の一環として行われた。
今回の研究では、以前産総研が発見した二次細胞壁(木質)の形成を制御するNST転写因子群の遺伝子破壊株であるシロイヌナズナnst1 nst3二重変異体をベースとなる実験材料として用いた。nst1 nst3二重変異体は2つのNST転写因子が破壊された植物で、茎の繊維細胞に二次細胞壁(木質)を形成できないので非常に折れやすく直立できない。二次細胞壁(木質)は、一般に陸上植物の乾燥重量の半分以上を占めることから、これを作っていないnst1 nst3二重変異体は外壁だけあって中身が空洞の「空っぽの容器」のようなものである。そこで、この「空っぽの容器」に、天然の二次細胞壁(木質)とは異なり産業利用に適した細胞壁を蓄積させることを目指した。
このnst1 nst3二重変異体に、約1800あるシロイヌナズナの全転写因子の遺伝子の中から、代表的な約300の転写因子を選んで導入した。導入した全ての植物を解析したところ、ERF転写因子遺伝子ファミリーの一部のサブファミリー(クラスIIId、IIIe)が導入された植物では、繊維細胞にリグニンを持たない細胞壁が蓄積していた(図1)。肥大した細胞壁の酵素糖化性を解析したところ、野生株と同等の細胞壁量(バイオマス量)を持ちながら2倍程度酵素糖化しやすいことがわかった(図2)。
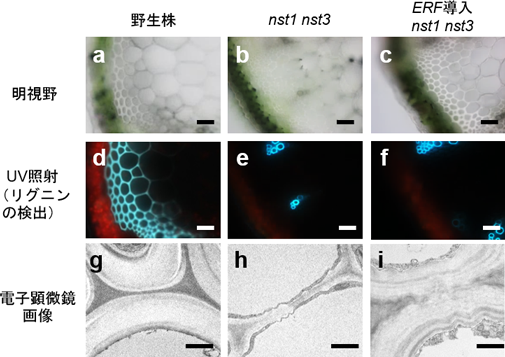 |
図1 クラスIIId、IIIe ERF転写因子群はリグニンのない細胞壁の蓄積を引き起こす
(スケールバー: 50 µm (a-f)、1 µm (g-i)) |
(a-c)シロイヌナズナの茎の切片の明視野観察像。
(d-f)紫外線(UV)照射によるリグニンの検出。野生株(d)ではリグニンの自家蛍光(水色)が細胞壁肥厚部位にはっきり見られるが、nst1 nst3二重変異体(e)やそこにクラスIIId、IIIe ERF転写因子を導入した植物(f)では道管にのみリグニンの自家蛍光が検出できる。
(g-i)繊維細胞の電子顕微鏡観察画像。 |
 |
|
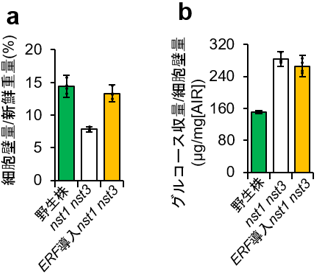
図2 ERF導入植物は糖化性の高い細胞壁を蓄積する |
|
ERF導入植物は野生株と同等の細胞壁量を持つにもかかわらず(a)、酵素糖化によるグルコース収量は野生株よりはるかに多い(b)。 |
導入した遺伝子の機能解析から、このクラスIIId、IIIe ERF転写因子群は植物の一次細胞壁の形成に関わる遺伝子の転写を制御していることがわかった。また、リグニンのない細胞壁の構成成分を分析したところ、一次細胞壁に特徴的なペクチンなどが多く含まれており、リグニンのない細胞壁は一次細胞壁が通常では起こりえないほど肥厚していたこともわかった(図1、図3)。
 |
|
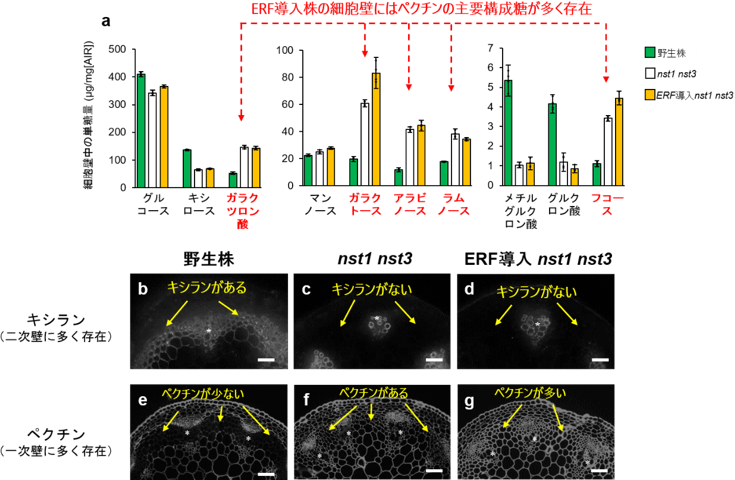
図3 クラスIIId、IIIe ERF転写因子が形成する細胞壁の糖組成は一次細胞壁に類似 |
(a)クラスIIId、IIIe ERF転写因子導入植物の細胞壁糖組成はほとんど一次細胞壁しか持たないnst1 nst3二重変異体に似ている。ガラクツロン酸やガラクトース、ラムノースなどペクチンの構成糖に富んでいる。
(b-g)茎の切片で二次細胞壁に多いキシランや一次細胞壁に多いペクチンを検出した。 |
一次細胞壁は通常リグニンを含まずセルロースやグルコースを取り出しやすいことから、今回の成果を活用して植物の一次細胞壁の割合を高めたり、二次細胞壁(木質)に一次細胞壁の特徴をブレンドしたりできれば、将来的に木質バイオマス由来の燃料や化成品の高効率生産に寄与すると期待される。
今後は、今回発見した一次細胞壁形成因子のより詳細な機能解明を進めつつ、肥厚させた一次細胞壁に通常木質に含まれる細胞壁成分を追加蓄積させるなどし、天然の木質に近い性質を持ちながら産業利用に適した木質を生産できるよう改良技術の開発を目指す。またポプラやユーカリなどの木本植物や、ソルガムやネピアグラス、スイッチグラスといった大型単子葉植物への適用を検討する。このような技術開発を継続することで石油資源のバイオマス資源への置換えを促進し、地球温暖化の抑制に貢献していく。