独立行政法人 産業技術総合研究所【理事長 野間口 有】(以下「産総研」という)生物プロセス研究部門【研究部門長 鎌形 洋一】合成生物工学研究グループ 宮崎 健太郎 研究グループ長、安武 義晃 主任研究員および国立大学法人 大阪大学大学院 情報科学研究科 北原 圭 研究員(前産総研特別研究員)らは、生物種に固有と考えられてきた16S rRNA遺伝子が異種生物由来のものにより置き換えることが可能であることを発見した。
リボソームは3種類のRNAと57種類のタンパク質から成る超分子複合体で、翻訳機能を担っている。その生理的役割の重要性と立体構造の複雑性から、進化的に極めて変異しにくい分子と考えられてきた。特にリボソーム30Sサブユニットに含まれる16S rRNAは、生物種固有の遺伝子としてバクテリアの系統分類学の指標となっている。今回、大腸菌の16S rRNA遺伝子欠損株に異種生物16S rRNAを導入して生育相補試験を行い、綱レベルで異なる遠縁の生物の16S rRNAであっても大腸菌内で機能することを実証した。この結果は、生物種固有であると考えられてきた16S rRNA遺伝子が種を超えて水平伝播しうる可能性を示唆し、従来の微生物の系統分類学の根本に疑問を投げかけるものである。
この成果の詳細は、2012年10月30日(日本時間)に米国の学術誌Proceedings of the National Academy of Sciences USA(米国科学アカデミー紀要)にオンライン掲載される。
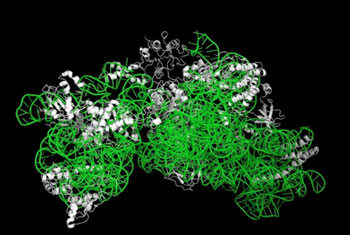 |
図1 リボソーム30Sサブユニットの立体構造
リボソーム30Sサブユニットは、16S rRNA(緑)と21のタンパク質(白)から構成される超分子複合体である。 |
リボソームは全生物がもつ細胞小器官であり、核酸にコードされた遺伝情報を機能(タンパク質)へと翻訳する。このような高次の生物機能を担うリボソームは、立体構造も複雑で、バクテリアでは3種のRNAと57種ものタンパク質から構成される。各構成成分は密接に相互作用しており、構成成分の遺伝子変異の多くは機能破壊(細胞死)を招くと考えられてきた。とくにRNAは立体構造上もリボソームの中心を占め、遺伝子の変異に対する感受性が高く、わずかな遺伝子変異がリボソーム機能の崩壊を招くと考えられてきた。実際、30Sサブユニットに含まれる16S rRNAは生物種に固有な分子として、長年、微生物の系統分類学の指標として使われている。
産総研では未知微生物の探索、ゲノム解析、有用遺伝子の探索、進化分子工学による遺伝子機能改変など、微生物機能の探索と利用に関する研究を幅広く行ってきた。最近では、リボソームの変異解析により大腸菌のリボソームに翻訳という本来の機能以外にも、リボヌクレアーゼの活性を阻害する機能が備わっていることを発見した(2011年11月25日 産総研プレス発表)。今回の成果も、リボソームの詳細な機能解析を行う中で得られたものである。
なお、今回の研究は文部科学省科学研究費補助金 新学術領域研究(領域名:動的・多要素な生体分子ネットワークを理解するための合成生物学の基盤構築)「翻訳システム改変による人工細胞創成」(平成24年度)、日本学術振興会 挑戦的萌芽研究「rRNAの置換変異によるリボソーム可塑性の研究」(平成24年度)、日本学術振興会 基盤研究(B)「メタゲノム遺伝子の網羅的発現を目指した大腸菌宿主の開発」(平成23~24年度)による支援を受けて行った。
産総研では、リボソーム30Sサブユニットに含まれる16S rRNAが異種生物由来のものに置き換えられるという予備的な知見をもっており、今回環境DNA(メタゲノム)から分離した16S rRNAを大腸菌に供給する手法により、網羅的・系統的な水平伝播実験を行った(図2)。
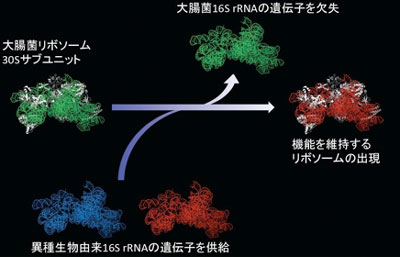 |
図2 リボソームの30Sサブユニットの立体構造と16S rRNA遺伝子の水平伝播実験
異種生物由来の16S rRNA(図中の赤色、青色)を大腸菌16S rRNA(緑色)と入れ替えて機能相補性に基づき選抜するかどうかを調べた。 |
環境DNAは環境中に含まれる微生物ゲノムの混合物で、極めて多様性に富んでいる。16S rRNA遺伝子の末端配列は微生物種間で相同性が高いため、共通のオリゴヌクレオチドプライマーを用いてさまざまな微生物由来の16S rRNA遺伝子をPCR(ポリメラーゼチェインリアクション)法により増幅することができる。こうして得たさまざまな生物種由来の16S rRNA遺伝子を大腸菌の16S rRNA欠損株に導入した。16S rRNA遺伝子は生育に必須であり、導入した16S rRNAが大腸菌で機能しないと細胞は生育しない。この性質を利用し、導入した16S rRNAの機能を判別した。その結果、大腸菌の16S rRNA遺伝子と綱のレベルで異なる微生物由来の16S rRNA遺伝子(配列相同性は80 %程度)でも大腸菌を生育させることができた。増殖の倍加時間は野生型の約35分と比較し遜色のないものから最大でも90分程度であった。
さらに、配列相同性の低い16S rRNAがいかにして複雑なリボソームに組み込まれるのか、配列の違いが構造・機能にどのように影響するのかを調べるために、得られた配列と大腸菌の該当する部分のそれぞれの二次構造を比較した。代表的な例として、ヘリックス21と呼ばれる領域について、配列の違いと二次構造を示す(図3)。塩基の配列が異なっていても二次構造が維持されれば、多くの場合16S rRNAの機能が維持されることが判明し、生物種固有と思われていた16S rRNAも、配列そのものではなく二次構造が機能を発揮するために重要であることがわかった。
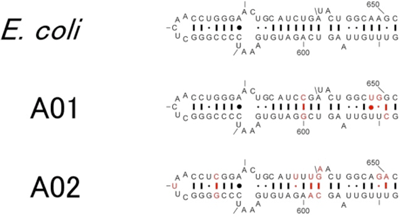 |
図3 大腸菌16S rRNAと異種生物由来の16S rRNAで大腸菌で機能するもののヘリックス21領域の構造
E.coliは大腸菌の16S rRNA遺伝子。A01、A02は、大腸菌16S rRNA遺伝子欠損株で機能した異種16S rRNA遺伝子の代表例。赤字は大腸菌と比較して変異している箇所。 |
生物種に固有と考えられてきた16S rRNA遺伝子が、綱のレベルで異なる生物種と相同性を示すという今回の発見は、16S rRNAには変異を受け入れる余地が十分にあるということを意味している。このことは16S rRNAの種特異性に基礎を置く従来の系統分類学に疑問を投げかけるものである。少数ではあるが、これまでの研究によって、ゲノム中に配列が大きく異なる複数の16S rRNAを含む微生物も報告されており、従来の系統分類学の見直しが求められる。
バクテリアで種を超えて16S rRNAの機能補完が起こりうることは、それを含むリボソーム自体が従来考えられていたよりもはるかに変化しやすい分子であることを意味している。これは、リボソームの積極的な機能改変へとつながる。リボソーム機能のわずかな違いは細胞内のタンパク質発現全体を変動させるため、リボソームの機能改変手法が細胞工学の新たな手法として発展する可能性がある。